Respiratory system & thermoregulation
The Respiratory System
The respiratory system is one of the major systems of the body. It has a number of very important functions including the provision of oxygen, the removal of carbon dioxide, the removal of excess heat (thermoregulation) and vocal communication. The respiratory system is a complex one and while there are some similarities with that of mammals, there are a number of quite significant differences.
Nasal cavity
The openings to the nasal cavity, the nares, lie at the point of the base of the comb on the top beak or mandible. The nasal cavity occupies a triangular shaped space between the nares and the margin of the eye and within the beak. Between the integument and the nasal cavity lie the lacrimal sinuses that empty into the cavity through the lateral wall.
The lateral wall of the cavity has three conchae, or projections, into the cavity:
- Anterior – of squamous epithelium – a single layer of flattened cells
- Medial – ciliated columnar epithelium – special cube shaped cells with cilia or hairs that trap foreign material
- Posterior – olfactory membrane – that gives the sense of smell
Smell, protection and reduction of electrolyte
The odour or smell of any material or object is a result of minute quantities of special chemicals that are detected by the olfactory membrane that in turn sends a signal via the olfactory nerve to the brain where it will be recognised for what it is. The epithelium of the nasal cavity is well endowed with mucosal glands which produce mucous that helps to keep foreign material from gaining entry to the body through the respiratory system. The nasal glands are small and located on either side. Their action is to supplement the action of the kidneys by reducing the electrolyte content of body fluids, especially when the level is higher than the kidneys can handle (electrolyte – common chemical salts e.g. sodium chloride).
Oropharynx (mouth and pharynx)
The oropharynx consists of the mouth and the pharynx that is located immediately behind it. The palate is part hard and part soft. The choanal opening (from the nasal cavity) is the cleft in the palate. The pharynx begins between the choanal opening and the common opening for the auditory tubes and extends to the rear to include that section of the oral cavity carrying the base of the tongue, the tip of which is located in the mouth.
Behind the base of the tongue is found the rima glottidis or opening into the larynx, which is sometimes called the cranial larynx. This opening is located in a conspicuous mound called the laryngeal prominence. The opening into the larynx is a median slit that is supported on each side by the arytenoid cartilages. These are special cartilages with a shape resembling the mouth of a jug or pitcher. It varies in length from approximately 8.5 mm in the female to 11 mm in the male. During gasping while the bird is in a state of respiratory distress it can open to a width of 7 to 9 mm. There are no vocal cords, epiglottis and thyroid cartilages that are normally found in mammals.
Trachea
This organ is a long tube with the function of moving the respiratory gases from the upper respiratory system to the organs of respiration – the lungs and air sacs or from the air sacs and lungs to the upper respiratory organs. The trachea in medium sized adults measures between 15 and 18 centimetres. It is held open permanently by 108 to 125 cartilaginous rings each one complete and lapping its neighbour. This arrangement prevents the trachea from collapsing or compressing but allows elongation and flexion. This means that while the trachea is flexible, under normal circumstances it will not collapse and prevent the free passage of air into/out of the lungs.
Most of the muscles in the region are involved in the movement of the head and neck and are not associated with respiration. The trachea is lined with muco-ciliary epithelium which is a special type of epithelium where the hair-like cilia move foreign materials, such as dust, up and out of the trachea. Numerous mucous secreting glands are also found in the tracheal lining.
Syrinx
The syrinx is the vocal organ of the fowl. It is located at the caudal end of the trachea and is suspended within the clavicular air sac. At rest, it is compressed laterally (at the sides). It consists of the pessulus, a wedge shaped cartilage located where the trachea divides into two to form the two bronchi plus the last four specialised cartilaginous rings at the bottom of the trachea. Below this section there are 4-7 thin, flexible syringeal cartilages which are fused at one end to the pessulus but which are free at the other. These are sometimes called the intermediate cartilages or syringeal ears.
At the syrinx the trachea forms two bronchi that then enter the lungs. After the syrinx, the cartilaginous rings of these bronchi are “C” shaped (i.e. an incomplete circle). The walls of the syrinx are two thin, vibrating membranes called the tympanic membranes. The bird makes sound by causing these membranes to vibrate. The tension of these membranes is controlled by muscles and the tension governs the sound produced.
Bronchi
The trachea divides at the syrinx into the left and right bronchi which are called the primary or mesobronchi. It is interesting to note that the combined cross sectional area of the bronchi is more than double that of the caudal end of the trachea from which they arise (30 mm2 v 12.5 mm2). This means that there is a significant reduction of air pressure and resistance to airflow in the bronchi than in the trachea. The cartilaginous rings of the bronchi extend from the syrinx to where the bronchi enter the lungs. Ciliated epithelium with numerous mucous glands lines the primary bronchi.
On entering the lungs, the primary bronchi divide to form four series of secondary bronchi and these, in turn, divide again to form numerous anastomising tertiary bronchi or parabronchi. The secondary and tertiary bronchi are lined with squamous epithelium and not the ciliated epithelium of the primary bronchi. The tertiary bronchi are arranged in layers. They are not blind ending, but join others that in turn lead back to the secondary and primary bronchi. Ultimately, the bronchial system is continuous.
Gas exchange
Leading off from the bronchi in the lungs are a large number of extremely small air capillaries (ducts) that are interlocked with the capillaries of the lung circulatory system. These interlocked capillaries are the lungs’ gas exchange system and are very thin which accommodates gaseous exchange. The blood/gas barrier in this area consists of whatever cell layers separate the two systems – the blood circulatory system and the air supply system of the lungs. These layers are:
- Single cell epithelial wall of the air capillary
- Base membrane one cell thick
- Single cell epithelial wall of the blood capillary
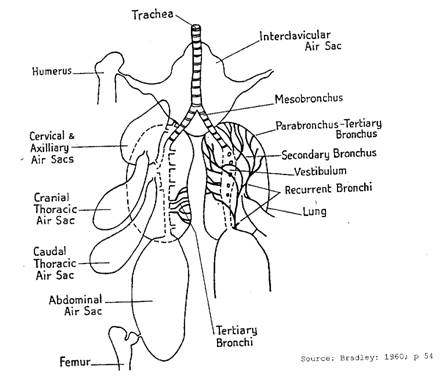
Air sacs and lungs
Lungs
The avian lung is a flattened structure that occupies the roof of the cranial or head end of the coelom. Fowls do not have a diaphragm that separates the peritoneal from the pericardial/pleural (thoracic) cavities. While on a body weight basis the volume of the avian lung is only one tenth of that of mammals of equal body size, it weighs significantly more (1.62 g/100 g to 1.38 g/100 g body weight). This indicates that the avian lung is very much denser than that of mammals.
The surfaces adjacent to the ribs are very indented and the ribs lie within the grooves that are formed such that approximately one third of the lungs lie between the ribs. The bright pink lungs are formed at the end of the bronchi and their terminal structures are called the air vesicles or sacs. Unlike other species, the avian lungs have very little elasticity.
Air sacs
The air sacs are very thin walled extensions of the bronchi that could be likened to balloons. Some of these sacs also connect to many of the larger long bones to form the pneumatic bones. This makes the bones lighter and is a distinct advantage in flight. There are nine air sacs in the domestic fowl. These are:
- Single clavicular air sac
- Pair of cervical air sacs
- Pair of cranial thoracic air sacs
- Pair of caudal thoracic air sacs
- Pair of abdominal air sacs
The clavicular sac is in the cranial end of the coelom. It has one median plus a pair of lateral chambers. The median chamber connects with the sternum (to which the keel bone is attached), coracoid bone and various ribs. The lateral chambers connect with the humerus. The syrinx is suspended in this sac. The cervical air sacs are located in the dorsal (towards the vertebrae) position at the front end of the coelom. They are connected to the cervical and thoracic vertebrae and the first two ribs. The cranial thoracic air sacs lie in the dorsolateral region (to the side towards the vertebrae) of the thoracic cavity.
The caudal thoracic air sacs, as the name implies, are located to the rear of the thoracic cavity. The abdominal air sacs extend from the rear of the lung to the region of the cloaca on each side and are quite large. They are attached to the wall of the abdominal cavity on one side and otherwise fill the spaces among the viscera. They connect with the sacrum and pelvic bones. There is an element of doubt as to whether they connect with the bones of the legs.
It is very difficult to see the air sacs in a bird because of the thin, transparent nature of the walls. Except for the abdominal sacs that connect directly with the primary bronchi, the air sacs connect with secondary and tertiary bronchi. It is believed the walls are lined with simple squamous epithelium although there is some ciliated columnar epithelium in some locations. There are very few blood vessels present which indicates that these sacs play no part in the gaseous exchange function of the respiratory system. In addition, the few blood vessels that do supply nutrients are part of the systemic system and not of the pulmonary circulatory system. There are very few nerves and muscles as well.
The air sacs provide a very high volume of air for use by the lungs on inspiration and expiration. Thus the lungs, aided by the air sac system, supply the oxygen necessary to power flight. The air sacs and hollow bones lighten the bird’s body which enhances its ability to fly. In addition, the air sac system permits the bird to change its centre of gravity, which also aids flight.
However, the non-flying ancestors of birds already possessed an air sac assisted breathing system, which possibly arose from a one-way lung system of a common ancestor of crocodilians and early dinosaurs. It may have been this hyper-efficient breathing system that provided an advantage to those ancestral archosaurs over the ancestors of mammals during a low-oxygen period of the early Triassic following the Permian mass-extinction event. In any case, some modern birds are able to fly comfortably in the cold, thin air above 9,000 metres.
The mechanics of respiration
There are two major actions associated with respiration – inspiration and expiration (inspiration – the drawing in of the air; expiration – the expulsion of air from the system). Respiration is a mechanical response by a number of muscle complexes to chemical and nerve controls as a result of situations that develop in the body:
- Oxygen low situation – when the body is in need of oxygen
- Carbon dioxide high situation – when the body needs to reduce the amount of carbon dioxide
- Thermoregulation demand – when the heat energy situation requires a response either to conserve or to remove it from the body
- Vocalisation – when the bird has a need to communicate by voice
Muscle complexes
The muscle complexes are in two groups. One group is associated with the inspiration and the other with expiration. While there are a number of muscles involved, there are two that are more important. These are the costisternalis pars major and the external intercostal muscle. If the operation of these is prevented respiration will cease, but if they remain functional and all others are non-functional, respiration continues at a reduced rate. Fowls do not have a diaphragm to divide the thoracic from the abdominal cavity. Respiration is the result of movement of the body external wall that in turn causes an increase or decrease in the volume of the cavity. These changes in volume cause changes in pressure inside of the lungs and this causes air to either enter or leave the system. Increasing the volume reduces the pressure and results in inspiration; decreasing the volume causes an increase in pressure and results in expiration.
Movement of the body wall
Stimulus of the muscles associated with inspiration will cause these muscles to contract, thus drawing the ribs forward. This pulls the sternum downwards and in effect increases the size of the cavity transversely and from front to back. The increase in size and volume lowers the pressure in the system and results in air from outside being drawn into the system to equalise the pressure. In other words, inspiration occurs as the air moves into the lungs and air sacs.
Inspiration
During inspiration the lungs are stretched by the action of the ribs (undergo distension). Approximately one third of the lungs lie between the rib bones and any movement of the ribs also affects the lungs. During inspiration the pressure in the system varies from high at the front (cranially) to low at the back (caudally) of the respiratory system. On the completion of inspiration the pressure will be generally equalised throughout the system. This occurs as the inspired air reaches the extremities of the respiratory system (the air sacs). At this stage the system consisting of the trachea, bronchi, the lungs, the air sacs and the thoracic cavity will be in a state of equilibrium.
Expiration
The above action is followed by the contraction of the muscles of expiration. This causes the ribs and sternum to return to their original position. This in turn will reduce the volume of the cavity. In this way the air pressure in the system will be increased above that of the outside atmosphere. As a consequence the air flows out of the system to the outside atmosphere until a situation of equilibrium is again reached.
Respiration cycle
While the operation of the lung and air sac system has been described above in a simple way, it does not operate quite so simply.
Bird lungs are radically different to the lungs of mammals. In mammals, the lung may be likened to closed bag. Fresh air is drawn in on inspiration for absorption of oxygen by small balloon-like alveoli connected to successively smaller branches of the bronchial tubes. Stale air is then expelled during expiration.
In birds, however, fresh air passes over the oxygen absorbing part of the lung, the parabronchi, on both inspiration and expiration. This means that birds, unlike mammals, are absorbing oxygen when they breathe in and when they breathe out. Using only the lungs, the caudal air sacs and cranial air sacs as a simple example, the bird’s lung and air sac system operates along the following lines:
- Inspiration 1: As fresh air enters the lungs from the trachea, some of it turns at the back of the lungs to go forward through the parabronchi for oxygen absorption and is then stored in the expanding cranial air sacs at the front of the body.
- Inspiration 2: The remainder of the fresh air reaching the back of the lungs is diverted directly into the expanding caudal air sacs at the back of the body.
- Expiration 1: As the cranial air sacs contract, stale air stored in them passes directly out of through the trachea.
- Expiration 2: As the caudal air sacks at the back of the body contract, fresh air stored in them now travels forward through the parabronchi (permitting the second cycle of oxygen absorption) and out through the trachea.
Gaseous exchange only takes place in the lungs and not in the air sacs. Thus, the bird respiration cycle uses both new air (referred to a neo or new respiration) and old air (referred to as paleo or old respiration). For a more detailed description, see the Bird Respiratory System.
Control of respiration
The control of respiration in birds, as in mammals, is exercised by chemical and nervous stimuli in the following way:
- Chemical control: appropriate receptors perceive the oxygen and carbon dioxide levels. When the oxygen level is too low or the carbon dioxide level too high, the respiratory system responds by increasing the rate of respiration. This would reflect the increased panting rate observed in the rarefied atmosphere at high altitude.
- Nervous control: there are three aspects of this part of control:
- Vagal reflexes – the vagus nerve system exercises influence over the respiratory system, heart and viscera. This is an involuntary control meaning that breathing cannot be stopped consciously for a long time. When the body needs to breathe it does so.
- Respiratory centre – located in the medulla of the brain, this centre coordinates respiratory processes. Not much is known about this centre, but it is generally believed that its function is more to do with thermoregulation than with the other functions of respiration.
- Thermal panting – this results from a build up of body heat usually from an increase in the environmental temperature or from additional activity (and increased metabolic rate).
Thermoregulation
As this term indicates, it is associated with the regulation of the temperature of the bird. Fowls are homeothermic animals. However, while every attempt is made to achieve a stable body temperature, there are times when, under extreme conditions, the birds’ temperature will vary up or down. When this variation is too great the bird is likely to die. The normal adult body temperature is in the range of 41-41.7oC and at approximately 46oC, death will occur.
Fat and heat loss
The layer of fat usually found under the skin plus the coat of feathers provide very good protection from low temperature and it is unlikely that birds other than young chickens will die of hypothermia. For a constant deep body temperature to be maintained, heat production must equal heat loss. Heat is produced by metabolic processes or may be gained from the environment when environmental temperature is above bird temperature. Birds lose heat to the atmosphere when the environmental temperature is below the birds body temperature, and when temperatures are approaching the birds’ body temperature if the relative humidity is low.
Heat is lost by the bird as sensible heat directly to the atmosphere when the temperature gradient is sufficiently great and as insensible heat by the evaporation of water from the respiratory system and skin when the temperature gradient is less but relative humidity is low. At high temperature the birds increase their respiration rate to increase the amount of air passing through the respiratory system to increase the cooling by evaporation. This panting also involves gular flutter which is the rapid movement of the upper throat tissues to increase evaporation. The movement of air around the body of the bird will assist in removing heat from the bird as sensible heat and as insensible heat.
High respiratory rate and eggshell formation
An increase in the respiratory rate at high temperatures will increase the loss of carbon dioxide from the body. While carbon dioxide is removed from the body because of its toxicity, a certain amount is used in the formation of eggshells. Eggshell is essentially calcium plus carbon dioxide that forms calcium carbonate. At higher environmental temperatures, layers often lay eggs with weaker shells because their panting to cool themselves removes more carbon dioxide from the body.
Summary of respiratory system and thermoregulation
An understanding of the structure and the functions of the respiratory system are very important if the farm manager is to maximise flock performance. Not only is the respiratory system the means of gaseous exchange for the birds, but also it plays an important role in the maintenance of a constant deep body temperature. Knowledge of how this is achieved is also important if the manager is to minimise losses from the effects of excess heat such as stock deaths and from lower growth or egg production and poor shell quality.
Further information
- Dingle, JG (1991) Module 3: Nutrient Procurement and Processing, Study Book: Poultry Husbandry 1, DEC, UCSQ, Toowoomba, Australia.
- Dingle, JG (1990) Module 4: Gas Exchange and Thermoregulation, Study Book: Poultry Husbandry 1, DEC, UCSQ, Toowoomba, Australia.
You May Also Like


The Poultry Hub Australia profoundly acknowledges and respects that its foundations, both people and facilities, are established on land rich in the history and traditions of the world’s oldest living culture. PHA values and respects Indigenous knowledge, understanding its importance in our shared history. We acknowledge the strength, resilience, and contributions of the Aboriginal community, we pay our tributes to the Aboriginal Elders – those who guided us in the past, those who lead us today, and those who will enlighten our paths in the future.